Tipos:
1. Mutaciones génicas o moleculares: afectan a la secuencia de bases del ADN.
TIPOS DE MUTACIONES GÉNICAS O MOLECULARES
Afectan a la secuencia de nucleótidos del gen, siendo invisibles con el microscopio. Dan lugar a nuevos alelos. Un ejemplo es la osteogénesis imperfecta o de los huesos de cristal, debida a una mutación génica en el gen del colágeno tipo I.
MUTACIONES POR SUSTITUCIÓN DE BASES
- TRANSICIONES: A por G o C por T, o viceversa (Púrica por púrica o pirimidínica por pirimidínica).
- TRANSVERSIONES: A o G por C o por T o viceversa (en los huesos de cristal una mutación puede ser la sustitución de G por T). Púrica por pirimidínica o viceversa.
Provocan la alteración de un sólo triplete de un gen. Puede provocar que se altere la función de la proteína o no (en el 1º caso, produce un cambio fenotípico, en el 2º, no)., dependiendo de si ha afectado a algún aa o no, y al lugar que ocupa el aa en la proteína. Las transiciones son más abundantes, pues provocan una alteración menor en la doble hélice, de modo que pasan más desapercibidas que las transversiones a los sistemas de reparación del ADN:
MUTACIONES POR PÉRDIDA O INSERCIÓN DE NUCLEÓTIDOS
- DELECIONES: se pierde un nucleótido (o más).
- INSERCIONES o ADICIONES: se añade un nucleótido (o más).
En ambos casos, además de alterarse la longitud de la proteína, se corre la pauta de lectura y a partir de ese punto (al no ser que se hayan eliminado o añadido 3) toda la proteína va a estar alterada.
CLASIFICACIÓN DE LAS MUTACIONES GÉNICAS POR SUS EFECTOS FENOTÍPICOS
1. SILENCIOSAS (o sinónimas)
El cambio de una base origina un aa sinónimo. Eso suele ocurrir por balanceo (cambio en el 3º nucleótido), aunque en el caso de la Leu, cambia también el 1º nucleótido.
2. NO SILENCIOSAS
2.1. CON CAMBIO DE SENTIDO. Ej., en la anemia falciforme, cuando el Glu de la posición 6 de la cadena beta de la hemoglobina ha mutado por una Val, resultado de una transversión en la que se ha cambiado T por A en el gen de la globina beta, situado en el cromosoma 11, dando una enfermedad autosómica recesiva (sólo los genotipos aa u homocigóticos recesivos desarrollan la enfermedad, mientras que los Aa o heterocigotos presentan resistencia a la malaria, por lo que el alelo a ha sufrido una presión selectiva en las zonas tropicales de África donde es endémica la malaria o paludismo). El cambio es más pronunciado cuando se cambia un aa polar por uno apolar, como en este caso, o viceversa. En este caso, la hemoglobina resultante, la hemoglobina S, forma unos polímeros anormales por interacciones hidrofóbicas entre la Val y otros aa apolares, y se deforma el eritrocito en forma de hoz, cuando no está transportando el oxígeno, entorpeciendo la circulación sanguínea. Afecta al 0,4% de la población afroamericana (aa) siendo el 8% portadores (Aa).
2.2. SIN SENTIDO. El nuevo codón es alguno de los tres codones sin sentido o de terminación (FIN), originando proteínas más cortas en su extremo c-terminal. Ejs.: fibrosis quística (enfermedad autosómica recesiva que afecta a los pulmones), distrofia muscular de Duchenne (miopatía recesiva ligada a X, el gen de la distrofina es el mayor gen conocido, con 2,3 Mb y 79 exones) y beta-talasemia (un tipo de anemia).
2.3. CON GANANCIA DE SENTIDO. Es lo inverso del caso anterior: un codón sin sentido muta a uno que significa un aa, dando proteínas más largas en su extremo C-terminal. Ej: la alfa-talasemia, en la que la alfa-hemoglobina es 30 aa más larga. La alfa-talasemia, como la anemia falciforme, produce ventaja frente a la malaria en los heterocigotos, de ahí su prevalencia en Sicilia y Cerdeña, donde un 10% de la población es portadora, por ser regiones donde había malaria).
3. ADICIONES O DELECIONES.
3.1. SIN CAMBIO DE PAUTA DE LECTURA. Si son en múltiplo de tres nucleótidos, no cambian la pauta. No son muy graves generalmente.
3.2. CON CAMBIO DE LA PAUTA DE LECTURA. A partir del punto mutado, toda la secuencia es incorrecta, a veces dando proteínas también más cortas, por lo que la mutación tiene consecuencias fatales al dar proteínas no funcionales.
2. Mutaciones cromosómicas: afectan a la estructura de los cromosomas.
a) inversiones: un fragmento de cromosoma se invierte, con lo que pueden inactivarse genes. Pueden disminuir la eficacia biológica (nº de descendientes) y, por tanto, se pueden eliminar por selección natural. Sin embargo, en ciertos casos pueden ser ventajosas y aumentar la eficacia biológica. Las inversiones son de gran importancia evolutiva ya que pueden ser un mecanismo de aislamiento reproductivo debido a la semiesterilidad del híbrido y al hecho de no existir recombinación en el segmento invertido. Todos los genes que se encuentran en el segmento invertido se transmiten siempre juntos y en ese orden, es como si formaran un grupo de ligamiento o un supergen que no sufre alteraciones por recombinación. Algunas inversiones cromosómicas se asocian a enfermedades en humanos como la Hemofilia A, un mayor riesgo de enfermedades neurodegenerativas o autoinmunes, así como con trastornos mentales. Sin embargo, no todas las inversiones se relacionan con enfermedad, existen algunas como una muy común entre la población europea que aumenta la fertilidad de sus portadores y por eso ha sido seleccionada positivamente en la población europea.
b) deleciones: un fragmento de ADN se elimina, con lo que pueden eliminarse genes. Suelen ser letales, al eliminarse información genética esencial, por lo que sufren una selección negativa (son eliminadas por selección natural).
c) duplicaciones: un fragmento de ADN se duplica, con lo que pueden aparecer genes por duplicado, p.ej. así pueden aparecer varias copias de genes fundamentales en la alimentación como ocurrió en la evolución de las poblaciones paleolíticas (cazadores-recolectores) a neolíticas. En las poblaciones agrícolas se seleccionaron mutaciones que incluían varias copias de amilasa, para digerir mejor los alimentos cultivados amiláceos, como las legumbres y cereales. en las poblaciones ganaderas, se seleccionaron mutaciones que incluían varias copias de lactasa, para digerir mejor la lactosa de la leche. Esto se comprueba, p.ej, con las poblaciones de Oriente Medio y Europa (agrícolas y ganaderas), que digieren sin dificultad el almidón y la lactosa, frente a las poblaciones de Extremo Oriente, que no digieren la lactosa en estado adulto o frente a las tribus amazónicas o de Nueva Guinea, que continúan un estado cazador-recolector.
d) transposiciones: un fragmento de ADN (transposón, elemento genético transponible, o "ADN saltarín") salta a otro lugar del genoma (el propio transposón o una copia), pudiendo provocar mutaciones en el lugar donde se insertan. Así, p.ej, surgen mazorcas de maíz con granos de diferentes colores, al haberse desactivado ciertos genes relacionados con el color. Fueron descubiertos por Barbara McClintock, por lo que recibió el Nobel. Su origen es incierto, tal vez proceden de antiguas secuencias víricas insertadas en el genoma eucariótico a lo largo de la evolución. De hecho, muchos son de origen retroviral y se transponen mediante una retrotranscriptasa. En ciertos casos, los fenotipos resultantes pueden ser ventajosos y ser seleccionados por selección Natural o Artificial (Mejora Genética Vegetal):

f) Translocaciones: un segmento de un cromosoma pasa a otro cromosoma de otra pareja. Hay un tipo de translocación en la que se intercambian segmentos entre cromosomas de diferentes parejas. Frecuentemente provocan enfermedades genéticas, disminuyendo la eficacia biológica, por lo que tienden a eliminarse por selección Natural. Un tipo de translocación produce un tipod e síndrome de Down (el Down familiar).
3. 3. Mutaciones genómicas: afectan al número de cromosomas
I. Aneuploidías
a) Que afectan a los autosomas (ej. Down).
Son las que afectan al nº de cromosomas, siendo detectables con el m.o. y pueden ser:
Aneupoloidías
Son cambios en los cromosomas por ganancia (trisomía) o pérdida de uno de ellos (monosomía).
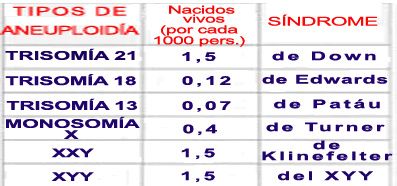
Se correlaciona con la edad de la madre (parece que relacionado con el tiempo que permanece la ovogénesis parada en diplotene, lo que puede hacer que el huso meiótico no se forme bien):
| Edad materna | Probabilidad de aneuploidía |
|---|
| 30 | 0,26% |
| 35 | 0,57% |
| 40 | 1,59% |
| 45 | 5,26% |
En casos excepcionales puede haber mujeres tetrasómicas XXXX y, más excepcionalmente, pentasómicas XXXXX. Dado que por cada X extra, disminuye el CI en, al menos 10, puntos, las últimas tienen un CI de alrededor de 65 (deficiencia mental).
- que afectan a los autosomas:
1. Síndrome de Down


 Fuente: «Down Syndrome Risk by Maternal Age-semilog» de Delphi234 - Trabajo propio. Disponible bajo la licencia CC0 vía Wikimedia Commons - http://commons.wikimedia.org/wiki/File:Down_Syndrome_Risk_by_Maternal_Age-semilog.svg#/media/File:Down_Syndrome_Risk_by_Maternal_Age-semilog.svg
Fuente: «Down Syndrome Risk by Maternal Age-semilog» de Delphi234 - Trabajo propio. Disponible bajo la licencia CC0 vía Wikimedia Commons - http://commons.wikimedia.org/wiki/File:Down_Syndrome_Risk_by_Maternal_Age-semilog.svg#/media/File:Down_Syndrome_Risk_by_Maternal_Age-semilog.svg
Se relaciona estadísticamente con una edad de la madre superior a los 35 años. Las personas con este síndrome tienen problemas cardíacos, digestivos y endocrinos, probablemente por el exceso de proteínas codificadas por el cromosoma 21.
Los varones con el síndrome son estériles (sólo se han reportado 3 casos en todo el mundo de varones fértiles). No así las mujeres, que pueden tener hijos con el síndrome e hijos sanos. La causa de la trisomía en un 15% se achaca al espermatozoide y en un 85% al óvulo.
2. Síndrome de Edwards (trisomía18): retraso mental.3. Síndrome de Patau: múltiples malformaciones.La mayoría de los casos de síndrome de Patau se deben a una trisomía del cromosoma 13 (consecuencia de una no disyunción meiótica, principalmente en el gameto materno). Aproximadamente un 20% de casos se deben a traslocaciones.
b) Que afectan a los cromosomas sexuales (ej. Klinefelter)
1. SÍNDROME DE TURNER: X0 (MONOSOMÍA)
Fue descubierta por el endocrinólogo estadounidense Henry Turner en 1938. Parece ser que la causa no es un fallo meiótico en la gametogénesis sino mitótico en la embriogénesis, por lo que estas mujeres suelen ser individuos-mosaico (XX/X0). Rendimiento escolar normal, con dificultades en las matemáticas y dibujo.
2. SÍNDROME DE KLINEFELTER: XXY (TRISOMÍA)

Causa hipogonadismo. En el 5% de los casos de SK pueden ser mosaicos cromosómicos con células incluso tetrasómicas (XXXY). Se ha insinuado que Carlos II de España, el hechizado, tenía el SK: Revista Médica de Chile, al menos en mosaico XY/XXY, como el de la foto del centro. Es la causa de hipogonadismo más frecuente y la aneuploidía de cromosomas sexuales también más frecuente, afectando a 1 entre 500 a 1.000 varones. Son estériles y presentan aspecto eunucoide. Durante la infancia puede pasar inadvertido, aunque a veces va acompañado de un ligero retraso mental.
La causa es la no disyunción meiótica:

Fuente: «XXY syndrome» de Silver Spoon - Trabajo propio. Disponible bajo la licencia CC BY-SA 3.0 vía Wikimedia Commons - https://commons.wikimedia.org/wiki/File:XXY_syndrome.svg#/media/File:XXY_syndrome.svg
3. TRIPLE X O SUPERHEMBRA: XXX (TRISOMÍA)
Se trata de individuos femeninos con órganos sexuales atrofiados, fertilidad limitada y bajo coeficiente intelectual. Su prevalencia es de aproximadamente 1 de cada 1.500 niñas. Los padres o las niñas afectadas probablemente no lleguen a darse cuenta de la presencia de esta enfermedad, a menos que se sometan a los exámenes médicos pertinentes. Suelen presenta retraso mental ("borderline"), aunque algunas han llegado a la universidad.
4. DOBLE Y (SUPERMACHO): XYY (TRISOMÍA)

La relación de esta enfermedad con comportamientos antisociales es polémica. Su desarrollo sexual es normal y suelen ser fértiles. Se ha reportado que su CI es 10 o 15 puntos inferior al de sus hermanos. Su incidencia es de 1 cada 1.000 niños y no parece ser afectada por la edad de los padres.
II. Poliploidía
La poliploidía se puede inducir mediante el uso de la colchicina que para la división celular antes de que se produzca la separación cromosómica. Las ventajas puede ser obtener semillas o frutos mayores. Un alopoliploide inducido, creado a finales del siglo XIX, es el Triticale, híbrido del trigo con el centeno.
Es un fenómeno frecuente en palntas cultivadas, como el trigo hexaploide, con 3 cereales ancestrales, los fresonres tetraploides y los plátanos triploides (estos últimos estériles pues para que la meiosis transcurra con normalidad el nº de cromosomas debe ser par).







 El ácido nitroso (HNO2) elimina grupos amino de la C, originando U., que se aparea con la A. Es un agente desaminante. De ahí, el carácter carcinógeno o mutagénico de los nitritos de los embutidos, añadidos en pequeñas cantidades para evitar la proliferación del clostridio del botulismo (bacteria que produce la toxina botulínica).
El ácido nitroso (HNO2) elimina grupos amino de la C, originando U., que se aparea con la A. Es un agente desaminante. De ahí, el carácter carcinógeno o mutagénico de los nitritos de los embutidos, añadidos en pequeñas cantidades para evitar la proliferación del clostridio del botulismo (bacteria que produce la toxina botulínica).







